
| 规格 | 价格 | 库存 | 数量 |
|---|---|---|---|
| 10 mM * 1 mL in DMSO |
|
||
| 1mg |
|
||
| 5mg |
|
||
| 10mg |
|
||
| 25mg |
|
||
| 50mg |
|
||
| 100mg |
|
||
| 250mg |
|
||
| 500mg |
|
||
| 1g |
|
||
| Other Sizes |
|

| 靶点 |
USP7 (IC50 = 0.19 μM); USP21(IC50 = 0.96 μM); Autophagy; NF-κB
|
|---|---|
| 体外研究 (In Vitro) |
体外活性:BAY 11-7082 完全且特异性地消除 NF-κB DNA 结合,下调 NF-κB 诱导细胞因子 IL-6 并诱导细胞凋亡。 BAY 11-7082 (< 8 μM) 能够以剂量依赖性方式有效抑制基础和 TNFα 刺激的 NFκB 荧光素酶活性。 BAY 11-7082 (8 μM) 强烈抑制 NCI-H1703 细胞的增殖速率。 Bay 11-7082 (5 μM) 快速有效地减少 HTLV-I 感染的 T 细胞系中 NF-kappaB 的 DNA 结合,并下调抗凋亡基因 Bcl-x(L) 的表达,而它具有对另一种转录因子 AP-1 的 DNA 结合几乎没有影响。 Bay 11-7082诱导的原代ATL细胞的凋亡比正常外周血单个核细胞更显着,并且这些细胞的凋亡还与NF-κB活性的下调有关。 Bay 11-7082 (5 μM) 选择性诱导 HTLV-I 感染的 T 细胞系凋亡,并与细胞周期蛋白 D1、细胞周期蛋白 D2 和 Bcl-xL 表达下调相关。 BAY 11-7082 (100 μM) 可防止 NMDA 引起的 p65 核转位以及 NMDA 诱导的小鼠海马切片中 NF-κB 结合的增加。 BAY 11-7082 可防止海马切片 CA1 区发生 NMDA 毒性,20 μM 时具有 40% 的神经保护作用,100 μM 时具有 70% 的神经保护作用。所有测试浓度的 BAY 11-7082 均显着抑制脂肪组织中的 NF-κB p65 DNA 结合活性,而在骨骼肌中,50 μM 和 100 μM 的 BAY 11-7082 显着抑制 NF-κB p65 DNA 结合活性。 BAY 11-7082 (100 μM) 可减少人脂肪组织和骨骼肌中的 IKK-β 蛋白。 BAY 11-7082 (100 μM) 显着降低脂肪组织中 TNF-α 的释放,而在所有测试浓度的 BAY 11-7082 下,IL-6 和 IL-8 的释放均受到显着抑制。 BAY 11-7082 (50 μM) 显着减少骨骼肌中 TNF-α、IL-6 和 IL-8 的释放。 BAY 11-7082 还被发现可以使 E2 结合酶 Ubc(泛素结合)13 和 UbcH7 以及 E3 连接酶 LUBAC(线性泛素组装复合物)失活,从而诱导 B 细胞淋巴瘤和白血病 T 细胞死亡激酶测定:将 UBE1 (0.17 μM) 溶解在 22.5 μL 20 mM Hepes(pH 7.5)中,含有 10 μM 泛素,与 1 μL DMSO 或 1 μL BAY 11-7082 的 DMSO 溶液一起在 21°C 下孵育 45 分钟。加入 2.5 μL 10 mM 醋酸镁和 0.2 mM ATP 溶液,30°C 孵育 10 分钟,加入 2.5 μL 10% (w/v) SDS 并加热 6 分钟终止反应在75°C。在不存在任何硫醇的情况下对样品进行 SDS/PAGE。将凝胶用考马斯速溶蓝染色 1 小时,然后用水洗涤脱色。以相同的方式将泛素加载到 E2 缀合酶上,不同之处在于,在与 BAY 11-7082 一起孵育之前将 UBE1 (0.17 μM) 与 Ubc13 (2.4 μM) 或 UbcH7 (2.9 μM) 混合。细胞测定:实验表明,Bay 11-7821 在有效抑制 IKK 的浓度下具有剧毒。它可以诱导MM细胞的细胞坏死。此外,Bay 11-7821 被证明通过诱导 B 细胞淋巴瘤和白血病 T 细胞死亡而具有抗炎能力。据报道,它还能抑制巨噬细胞中的 NALP3 炎性小体
|
| 体内研究 (In Vivo) |
异种移植模型证实了BAY 11-7082在体内诱导细胞凋亡和抑制生长的抗肿瘤作用[8]。
最后,建立人胃癌异种移植瘤模型,验证BAY 11-7082的体内抗肿瘤作用。TUNEL和免疫组化检测皮下肿瘤切片细胞凋亡和生长抑制[8]。 在体内短期局部应用BAY 11-7082防止暴露小鼠下咽粘膜酸性胆汁诱导的mRNA和miRNA致癌表型。https://pubmed.ncbi.nlm.nih.gov/29529473/ NF-kappaB是一种核转录因子,参与控制包括细胞存活在内的基本细胞功能。在该因子的众多靶基因中,有促凋亡基因和抗凋亡基因。为了评估NF-kappaB激活对兴奋性毒性损伤的贡献,我们分析了IkappaBalpha (IkappaBalpha)磷酸化阻断对成年小鼠海马切片中谷氨酸诱导的毒性的影响。通过免疫细胞化学和EMSA技术,我们发现(i)海马切片急性暴露于NMDA诱导NF-kappaB核易位,(ii) NMDA介导的NF-kappaB活化被IkappaBalpha磷酸化和降解抑制剂BAY 11-7082阻止,(iii) BAY 11-7082介导的NF-kappaB活化抑制与神经保护有关。[5] |
| 酶活实验 |
将 UBE1 (0.17 μM) 溶解在 22.5 μL 20 mM Hepes(pH 7.5)中,含有 10 μM 泛素,与 1 μL DMSO 或 1 μL BAY 11-7082 的 DMSO 溶液一起在 21°C 下孵育 45 分钟。加入 2.5 μL 10 mM 醋酸镁和 0.2 mM ATP 溶液,30°C 孵育 10 分钟,加入 2.5 μL 10% (w/v) SDS 并加热 6 分钟终止反应在75°C。在不存在任何硫醇的情况下对样品进行 SDS/PAGE。将凝胶用考马斯速溶蓝染色 1 小时,然后用水洗涤脱色。以相同的方式将泛素加载到 E2 缀合酶上,不同之处在于,在与 BAY 11-7082 一起孵育之前将 UBE1 (0.17 μM) 与 Ubc13 (2.4 μM) 或 UbcH7 (2.9 μM) 混合。
|
| 细胞实验 |
在 96 孔微量滴定板中,将 siRNA 转染到细胞中,然后将细胞在完全 NSCLC 培养基中培养 72 小时,并给予 12 小时 BAY 11-7082 处理。将细胞与[3H]胸苷一起孵育三个小时。使用自动细胞收集器将细胞收集到过滤器上后,通过 β-闪烁计数确定过滤器上的放射性。
此外,BAY 11-7082抑制NF-κB信号通路可抑制细胞增殖;这表明Rac1表达沉默导致的细胞增殖和迁移的丧失可能部分归因于NF-κB活性的丧失。[2] 人类t细胞白血病病毒I型(HTLV-I)是一种称为成人t细胞白血病(ATL)的侵袭性白血病的病原体。我们之前已经证明,所有被HTLV-I感染的t细胞系和来自ATL患者的原发性白血病细胞都表现出高活性的转录因子NF-kappaB。在本研究中,我们发现NF-kappaB抑制剂Bay 11-7082可诱导htlv -i感染的T细胞系凋亡,但对htlv -i阴性的T细胞凋亡作用微不足道。在htlv -i感染的t细胞株中,Bay 11-7082能快速有效地降低NF-kappaB的DNA结合,并下调NF-kappaB调控的抗凋亡基因Bcl-x(L)的表达,而对另一转录因子AP-1的DNA结合影响不大。虽然病毒蛋白Tax是NF-kappaB的激活剂,但Bay 11-7082诱导htlv -i感染细胞的凋亡与Tax的表达降低无关。此外,Bay 11-7082诱导的原代ATL细胞的凋亡比正常外周血单个核细胞的凋亡更为突出,并且这些细胞的凋亡也与NF-kappaB活性的下调有关。我们的研究结果表明,NF-kappaB在htlv -i感染的白血病细胞的发病和存活中起着至关重要的作用,是预防和治疗ATL的合适靶点。[4] |
| 动物实验 |
Male BALB/c nude mice.
2.5 & 5 mg/kg i.t. In this work, researchers topically exposed HM (C57Bl/6j wild-type) to a mixture of bile acids at pH 3.0 with and without BAY 11-7082 3 times/day for 7 days. They used immunofluorescence, Western blotting, immunohistochemistry, quantitative polymerase chain reaction, and polymerase chain reaction microarrays to identify NF-κB activation and its associated oncogenic mRNA and miRNA phenotypes, in murine hypopharyngeal cells in vitro and in murine HM in vivo. Results: Short-term exposure of HM to acidic bile is a potent stimulus accelerating the expression of NF-κB signaling (70 out of 84 genes) and oncogenic molecules. Topical application of BAY 11-7082 sufficiently blocks the effect of acidic bile. BAY 11-7082 eliminates NF-κB activation in regenerating basal cells of acidic bile-treated HM and prevents overexpression of molecules central to head and neck cancer, including bcl-2, STAT3, EGFR, TNF-α, and WNT5A. NF-κB inhibitor reverses the upregulated "oncomirs" miR-155 and miR-192 and the downregulated "tumor suppressors" miR-451a and miR-375 phenotypes in HM affected by acidic bile. Conclusion: There is novel evidence that acidic bile-induced NF-κB-related oncogenic mRNA and miRNA phenotypes are generated after short-term 7-day mucosal exposure and that topical mucosal application of BAY 11-7082 can prevent the acidic bile-induced molecular alterations associated with unregulated cell growth and proliferation of hypopharyngeal cells. https://pubmed.ncbi.nlm.nih.gov/29529473/ |
| 参考文献 |
[1]. Expert Opin Ther Targets . 2007 Feb;11(2):133-44. [2]. Cancer Biol Ther . 2012 Jun;13(8):647-56. [3]. J Med Chem . 2005 Sep 22;48(19):5966-79. [4]. Blood . 2002 Sep 1;100(5):1828-34. [5]. Neurosci Lett . 2005 Apr 4;377(3):147-51. [6]. Biochem J . 2013 May 1;451(3):427-37. |
| 其他信息 |
(E)-3-tosylacrylonitrile is a nitrile that is acrylonitrile in which the hydrogen located beta,trans to the cyano group is replaced by a tosyl group. It is an inhibitor of cytokine-induced IkappaB-alpha phosphorylation in cells. It has a role as a non-steroidal anti-inflammatory drug, an EC 2.7.11.10 (IkappaB kinase) inhibitor, an EC 3.1.3.48 (protein-tyrosine-phosphatase) inhibitor, a platelet aggregation inhibitor and an apoptosis inducer. It is a sulfone and a nitrile.
(E)-3-Tosylacrylonitrile has been reported in Aspergillus terreus with data available. The small GTPase Rac1 regulates many cellular processes, including cytoskeletal reorganization, cell migration, proliferation, and survival. Additionally, Rac1 plays a major role in activating NF-κB-mediated transcription. Both Rac1 and NF-κB regulate many properties of the malignant phenotype, including anchorage-independent proliferation and survival, metastasis, and angiogenesis. Despite these findings, the roles of Rac1and NF-κB in non-small cell lung carcinoma, a leading cause of cancer deaths, have not been thoroughly investigated. Here, we compared the effects of Rac1 siRNA to that of the Rac1 inhibitor NSC23766 on multiple features of the NSCLC malignant phenotype, including NF-κB activity. We show that the siRNA-mediated silencing of Rac1 in lung cancer cells results in decreased cell proliferation and migration. The decrease in proliferation was observed in both anchorage-dependent and anchorage-independent assays. Furthermore, cells with decreased Rac1 expression have a slowed progression through the G 1 phase of the cell cycle. These effects induced by Rac1 siRNA correlated with a decrease in NF-κB transcriptional activity. Additionally, inhibition of NF-κB signaling with BAY 11-7082 inhibited proliferation; indicating that the loss of cell proliferation and migration induced by the silencing of Rac1 expression may be attributed in part to loss of NF-κB activity. Interestingly, treatment with the Rac1 inhibitor NSC23766 strongly inhibits cell proliferation, cell cycle progression, and NF-κB activity in lung cancer cells, to an even greater extent than the inhibition induced by Rac1 siRNA. These findings indicate that Rac1 plays an important role in lung cancer cell proliferation and migration, most likely through its ability to promote NF-κB activity, and highlight Rac1 pathways as therapeutic targets for the treatment of lung cancer.[2] Human T-cell leukemia virus type I (HTLV-I) is the causative agent of an aggressive form of leukemia designated adult T-cell leukemia (ATL). We have previously demonstrated that all T-cell lines infected with HTLV-I and primary leukemic cells from ATL patients display constitutively high activity of transcription factor NF-kappaB. In this study we showed that Bay 11-7082, an inhibitor of NF-kappaB, induced apoptosis of HTLV-I-infected T-cell lines but only negligible apoptosis of HTLV-I-negative T cells. Bay 11-7082 rapidly and efficiently reduced the DNA binding of NF-kappaB in HTLV-I-infected T-cell lines and down-regulated the expression of the antiapoptotic gene, Bcl-x(L), regulated by NF-kappaB, whereas it had little effect on the DNA binding of another transcription factor, AP-1. Although the viral protein Tax is an activator of NF-kappaB, Bay 11-7082-induced apoptosis of HTLV-I-infected cells was not associated with reduced expression of Tax. Furthermore, Bay 11-7082- induced apoptosis of primary ATL cells was more prominent than that of normal peripheral blood mononuclear cells, and apoptosis of these cells was also associated with down-regulation of NF-kappaB activity. Our results indicate that NF-kappaB plays a crucial role in the pathogenesis and survival of HTLV-I-infected leukemic cells and that it is a suitable target for the prevention and treatment of ATL.[4] The compound BAY 11-7082 inhibits IκBα [inhibitor of NF-κB (nuclear factor κB)α] phosphorylation in cells and has been used to implicate the canonical IKKs (IκB kinases) and NF-κB in >350 publications. In the present study we report that BAY 11-7082 does not inhibit the IKKs, but suppresses their activation in LPS (lipopolysaccharide)-stimulated RAW macrophages and IL (interleukin)-1-stimulated IL-1R (IL-1 receptor) HEK (human embryonic kidney)-293 cells. BAY 11-7082 exerts these effects by inactivating the E2-conjugating enzymes Ubc (ubiquitin conjugating) 13 and UbcH7 and the E3 ligase LUBAC (linear ubiquitin assembly complex), thereby preventing the formation of Lys63-linked and linear polyubiquitin chains. BAY 11-7082 prevents ubiquitin conjugation to Ubc13 and UbcH7 by forming a covalent adduct with their reactive cysteine residues via Michael addition at the C3 atom of BAY 11-7082, followed by the release of 4-methylbenzene-sulfinic acid. BAY 11-7082 stimulated Lys48-linked polyubiquitin chain formation in cells and protected HIF1α (hypoxia-inducible factor 1α) from proteasomal degradation, suggesting that it inhibits the proteasome. The results of the present study indicate that the anti-inflammatory effects of BAY 11-7082, its ability to induce B-cell lymphoma and leukaemic T-cell death and to prevent the recruitment of proteins to sites of DNA damage are exerted via inhibition of components of the ubiquitin system and not by inhibiting NF-κB.[6] |
| 分子式 |
C10H9NO2S
|
|
|---|---|---|
| 分子量 |
207.25
|
|
| 精确质量 |
207.035
|
|
| 元素分析 |
C, 57.96; H, 4.38; N, 6.76; O, 15.44; S, 15.47
|
|
| CAS号 |
19542-67-7
|
|
| 相关CAS号 |
|
|
| PubChem CID |
5353431
|
|
| 外观&性状 |
White to off-white solid powder
|
|
| 密度 |
1.2±0.1 g/cm3
|
|
| 沸点 |
397.6±42.0 °C at 760 mmHg
|
|
| 熔点 |
133-135℃
|
|
| 闪点 |
194.3±27.9 °C
|
|
| 蒸汽压 |
0.0±0.9 mmHg at 25°C
|
|
| 折射率 |
1.557
|
|
| LogP |
1.28
|
|
| tPSA |
66.31
|
|
| 氢键供体(HBD)数目 |
0
|
|
| 氢键受体(HBA)数目 |
3
|
|
| 可旋转键数目(RBC) |
2
|
|
| 重原子数目 |
14
|
|
| 分子复杂度/Complexity |
347
|
|
| 定义原子立体中心数目 |
0
|
|
| SMILES |
S(/C=C/C#N)(C1C=CC(C)=CC=1)(=O)=O
|
|
| InChi Key |
DOEWDSDBFRHVAP-KRXBUXKQSA-N
|
|
| InChi Code |
InChI=1S/C10H9NO2S/c1-9-3-5-10(6-4-9)14(12,13)8-2-7-11/h2-6,8H,1H3/b8-2+
|
|
| 化学名 |
(E)-3-(4-methylphenyl)sulfonylprop-2-enenitrile
|
|
| 别名 |
BAY 11-7821; BAY-11-7821; BAY11-7821; bay 11-7082; 19542-67-7; (E)-3-Tosylacrylonitrile; (E)-3-(p-Toluenesulfonyl)acrylonitrile; Bay 11-7821; (E)-3-(4-Methylphenyl)sulfonylprop-2-enenitrile; BAY11-7082;BAY 11-7082; BAY11-7082; BAY 117082; BAY117082; BAY-117082; BAY-11-7082
|
|
| HS Tariff Code |
2934.99.9001
|
|
| 存储方式 |
Powder -20°C 3 years 4°C 2 years In solvent -80°C 6 months -20°C 1 month |
|
| 运输条件 |
Room temperature (This product is stable at ambient temperature for a few days during ordinary shipping and time spent in Customs)
|
| 溶解度 (体外实验) |
|
|||
|---|---|---|---|---|
| 溶解度 (体内实验) |
配方 1 中的溶解度: ≥ 2.5 mg/mL (12.06 mM) (饱和度未知) in 10% DMSO + 40% PEG300 + 5% Tween80 + 45% Saline (这些助溶剂从左到右依次添加,逐一添加), 澄清溶液。
例如,若需制备1 mL的工作液,可将100 μL 25.0 mg/mL澄清DMSO储备液加入到400 μL PEG300中,混匀;然后向上述溶液中加入50 μL Tween-80,混匀;加入450 μL生理盐水定容至1 mL。 *生理盐水的制备:将 0.9 g 氯化钠溶解在 100 mL ddH₂O中,得到澄清溶液。 配方 2 中的溶解度: ≥ 2.5 mg/mL (12.06 mM) (饱和度未知) in 10% DMSO + 90% Corn Oil (这些助溶剂从左到右依次添加,逐一添加), 澄清溶液。 例如,若需制备1 mL的工作液,可将 100 μL 25.0 mg/mL 澄清 DMSO 储备液添加到 900 μL 玉米油中并混合均匀。 View More
配方 3 中的溶解度: 30% PEG400+0.5% Tween80+5% propylene glycol: 15 mg/mL 1、请先配制澄清的储备液(如:用DMSO配置50 或 100 mg/mL母液(储备液)); 2、取适量母液,按从左到右的顺序依次添加助溶剂,澄清后再加入下一助溶剂。以 下列配方为例说明 (注意此配方只用于说明,并不一定代表此产品 的实际溶解配方): 10% DMSO → 40% PEG300 → 5% Tween-80 → 45% ddH2O (或 saline); 假设最终工作液的体积为 1 mL, 浓度为5 mg/mL: 取 100 μL 50 mg/mL 的澄清 DMSO 储备液加到 400 μL PEG300 中,混合均匀/澄清;向上述体系中加入50 μL Tween-80,混合均匀/澄清;然后继续加入450 μL ddH2O (或 saline)定容至 1 mL; 3、溶剂前显示的百分比是指该溶剂在最终溶液/工作液中的体积所占比例; 4、 如产品在配制过程中出现沉淀/析出,可通过加热(≤50℃)或超声的方式助溶; 5、为保证最佳实验结果,工作液请现配现用! 6、如不确定怎么将母液配置成体内动物实验的工作液,请查看说明书或联系我们; 7、 以上所有助溶剂都可在 Invivochem.cn网站购买。 |
| 制备储备液 | 1 mg | 5 mg | 10 mg | |
| 1 mM | 4.8251 mL | 24.1255 mL | 48.2509 mL | |
| 5 mM | 0.9650 mL | 4.8251 mL | 9.6502 mL | |
| 10 mM | 0.4825 mL | 2.4125 mL | 4.8251 mL |
1、根据实验需要选择合适的溶剂配制储备液 (母液):对于大多数产品,InvivoChem推荐用DMSO配置母液 (比如:5、10、20mM或者10、20、50 mg/mL浓度),个别水溶性高的产品可直接溶于水。产品在DMSO 、水或其他溶剂中的具体溶解度详见上”溶解度 (体外)”部分;
2、如果您找不到您想要的溶解度信息,或者很难将产品溶解在溶液中,请联系我们;
3、建议使用下列计算器进行相关计算(摩尔浓度计算器、稀释计算器、分子量计算器、重组计算器等);
4、母液配好之后,将其分装到常规用量,并储存在-20°C或-80°C,尽量减少反复冻融循环。
计算结果:
工作液浓度: mg/mL;
DMSO母液配制方法: mg 药物溶于 μL DMSO溶液(母液浓度 mg/mL)。如该浓度超过该批次药物DMSO溶解度,请首先与我们联系。
体内配方配制方法:取 μL DMSO母液,加入 μL PEG300,混匀澄清后加入μL Tween 80,混匀澄清后加入 μL ddH2O,混匀澄清。
(1) 请确保溶液澄清之后,再加入下一种溶剂 (助溶剂) 。可利用涡旋、超声或水浴加热等方法助溶;
(2) 一定要按顺序加入溶剂 (助溶剂) 。
BAY 11-7082 suppresses the activation of IKK family members and JNK.Biochem J. 2013 May 1;451(3):427-437. |
|---|
BAY 11-7082 and BAY 11-7085, but not other inhibitors of the activity and activation of the canonical IKK complex, induce HBL-1 cell death.
BAY 11-7082 forms a covalent adduct with Ubc13. |
BAY 11-7082 does not affect the LPS-stimulated autophosphorylation of IRAK4.
BAY 11-7082 suppresses the LPS- or IL-1-stimulated formation of K63-pUb chains and the DNA damage response.Biochem J. 2013 May 1;451(3):427-437. |
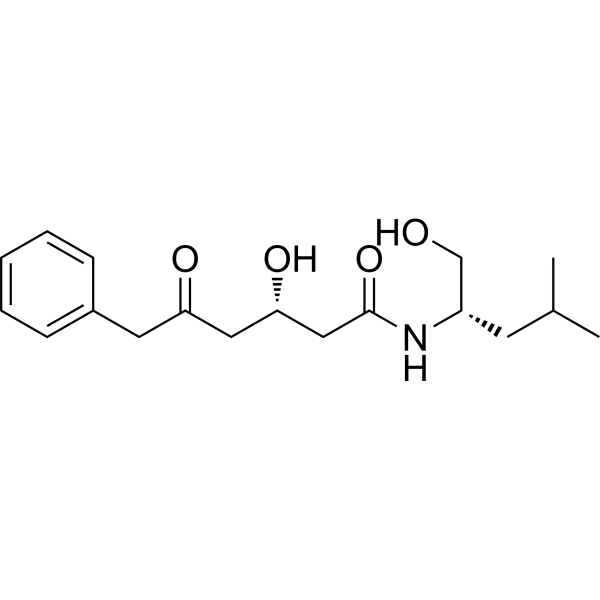 IKKβ-IN-3
IKKβ-IN-3
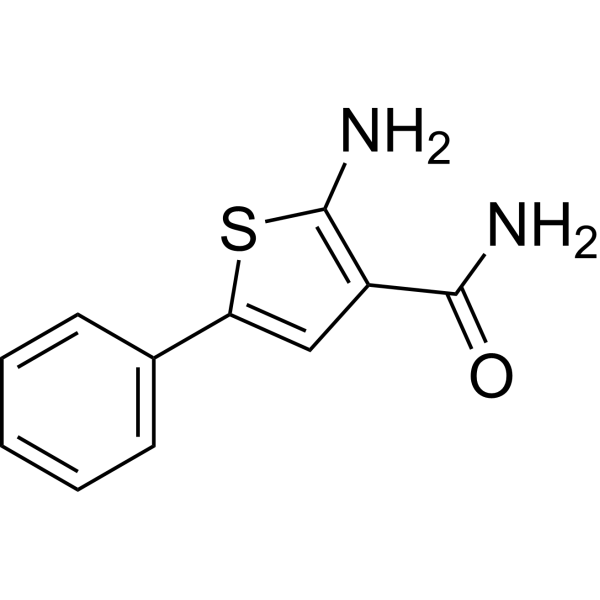 IKKβ-IN-4
IKKβ-IN-4
 Resolvin D1-d5 (RvD1-d5)
Resolvin D1-d5 (RvD1-d5)
 6-keto Prostaglandin F1α
6-keto Prostaglandin F1α
InvivoChem的所有产品仅用于作科学研究,不面向患者销售
Copyright 2020 InvivoChem LLC | All Rights Reserved 粤ICP备20063088号-1





























 COA
COA





